研究テーマ
慢性的なDNA損傷ストレスに対する耐性獲得の分子メカニズム
生物の遺伝情報を担うゲノムDNA は、電離放射線、紫外線、化学物質などの外的要因の他、細胞活動等によって発生する活性酸素などの内的要因によって絶えず損傷を受けている。これらに共通する点は、微量かつ慢性的にゲノム上のランダムな場所で発生しているということである。生物は、この慢性的な損傷ストレスに対して耐性を獲得することで地球環境に適応してきた一方で、このような長きに渡る損傷への暴露がヒトの発がんや老化と密接に関連していると考えられている。DNA損傷ストレスの防御機構には、DNA損傷修復機構の他に、細胞周期や遺伝子発現などを制御する損傷チェックポイント機構や、複製阻害の解消を行うDNA損傷トレランス機構などが知られており、実際の細胞内では、これらの機構が独立に機能するのではなく、DNA損傷ストレスに対して連携しながら、細胞の増殖とゲノムの安定性を維持する一つのシステムとして機能していると考えられる。当研究室では、自然環境レベルの紫外線照射下で酵母細胞を培養する装置を独自に開発し、慢性的なDNA損傷ストレス環境における細胞の損傷応答システムの解明と、これらのシステムの破綻が引き起こすゲノム不安定性について詳細に解析している。さらに、本研究成果をゲノム不安定性を伴うヒトの疾患の原因解明に役立てることで、それらの生物学的・医学的意義を統合的に理解することを目指している。
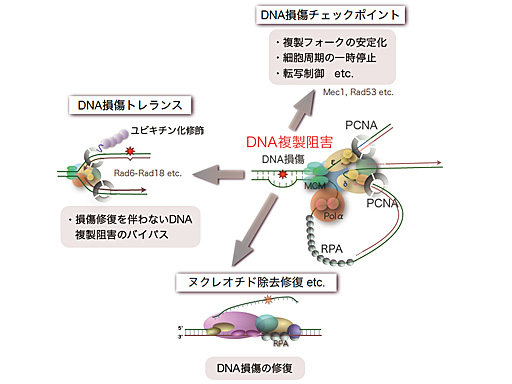
紫外線による複製阻害と損傷応答
DNA損傷トレランス経路の分子メカニズムの解明
DNA損傷によるDNA複製フォークの進行阻害は、ゲノム不安定性を引き起こす主要な原因である。複製フォークの進行阻害が起こった場合、複製の再開にDNA損傷の修復を待っていてはDNA複製の完了に膨大な時間が必要となってしまう。そのような事態を回避するために、DNA損傷を残したまま複製フォークが損傷部位を乗り越えるDNA損傷トレランスと呼ばれる機構が存在する。DNA損傷トレランスの制御には、ユビキチン化と呼ばれる翻訳後修飾が重要な役割を果たしており、DNA複製阻害が引き起こされると、DNA複製装置に含まれるPCNA(DNAクランプ)というタンパク質がユビキチン化修飾を受け、損傷トレランス経路を活性化することが知られている。さらに、この修飾にはユビキチンタンパク質が1つだけ結合したモノユビキチン化と複数のユビキチンがポリマー状に結合しているポリユビキチン化の2つの修飾形態が存在し、それぞれ別の損傷トレランス経路を活性化している。このユビキチン化修飾の制御機構や複製阻害部位のバイパス機構に関しては、未だ不明な点が多く残されており、現在詳細な解析を行っている。
DNA相同組換えの分子メカニズムの解明とその応用
DNA組換え反応は、バクテリアからヒトまで高度に保存された反応であり、体細胞、減数分裂細胞、免疫細胞、さらにはバイオテクノロジーの分野において様々な役割を果たしている。例えば、体細胞においては、電離放射線や変異源物質などによって生じるDNA鎖切断の修復のほか、DNA複製阻害の解消やテロメア構造の維持など、染色体の安定性維持に重要な役割を果たしている。免疫細胞や減数分裂細胞では、それぞれ抗原受容遺伝子の再編成や相同染色体間の組換えに関与しており、これらはいずれも多様性の獲得に貢献している。このように、DNA組換えは、遺伝情報の本体であるゲノムに対して「安定と変化」をもたらす両面の働きを持っている。私たちの研究室では、出芽酵母及び大腸菌をモデル生物として用いて、DNA相同組換えの分子メカニズムを詳細に解析し、ゲノム安定性維持機構の解明を目指している。
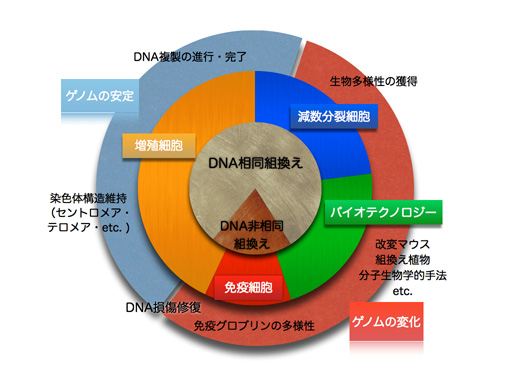
DNA相同組換えの様々な役割
DNA相同組換えの制御メカニズムの解明
DNA相同組換え反応は、DNAヘリカーゼ、ヌクレアーゼ、DNA鎖交換タンパク質の他、様々なアクセサリータンパク質が関与しており、その反応は、ATP加水分解のエネルギーを利用する多段階からなる機械的反応である。その中でもDNAヘリカーゼなどのDNAモータータンパクは、組換え制御因子として中心的な役割を果たしている。我々の研究室では、Sgs1、Srs2、Mgs1などのDNA二重鎖を巻き戻す(巻き直す)活性を持つタンパク質に着目し、これらがDNA相同組換え制御において果たす役割を解析している。特に、出芽酵母Sgs1タンパク質は、大腸菌からヒトまで高度に保存され、ヒトのSgs1ホモログであるBLM及びWRNは、それぞれ高発癌性のブルーム症候群や早期老化が現れるウェルナー症候群の原因遺伝子産物であることから、これらの分子メカニズムの解明が発がんや老化の分子基盤の解明につながるものとして期待されている。
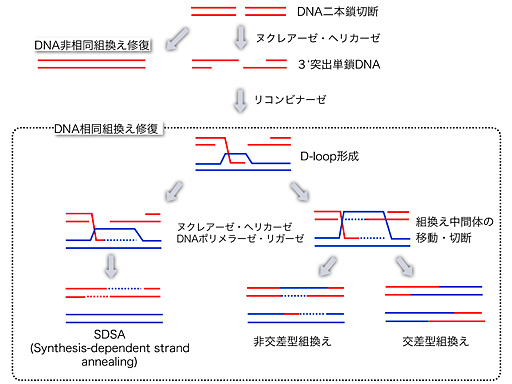
DNA相同組換え経路
活動実績
主な論文
Keyamura, K., Arai, K., and Hishida, T.
Srs2 and Mus81-Mms4 prevent accumulation of toxic inter-homolog recombination intermediates. PLoS Genet. 12(7), (2016).
Keyamura, K., Sakaguchi, C., Kubota, Y., Niki, H. and Hishida, T.
RecA recruits SMC-like RecN to DNA double-strand breaks. J. Biol. Chem. 288, 29229-37 (2013).
Haruta, N., Kubota, Y. and Hishida, T.
Chronic low-dose ultraviolet induced mutagenesis in nucleotide excision repair-deficient cells. Nucleic Acids Res. 40, 8406-8415 (2012).
Hishida, T., Hirade, Y., Haruta, N., Kubota, Y. and Iwasaki, H.
Srs2 controls DNA damage checkpoint responses via two distinct homologous recombination functions in Saccharomyces cerevisiae. Mol. Cell. Biol. 30: 4840-4850 (2010).
Hishida, T., Kubota, Y., Carr, A. M. and Iwasaki, H.
RAD6-RAD18-RAD5 pathway-dependent tolerance to chronic low-dose UV light. Nature 457: 612-615 (2009).
Hishida, T., Ohya, T., Yoshino Kubota, Y., Kamada Y. and Shinagawa, H.
Functional and physical interaction of yeast Mgs1 with PCNA: impact on RAD6-dependent DNA damage tolerance. Mol. Cell. Biol. 26: 5509-5517 (2006).
Hishida, T., Han, Y-W., Shibata, T., Kubota, Y., Ishino, Y., Iwasaki , H. and Shinagawa, H.
Role of the Escherichia coli RecQ DNA helicase in SOS signaling and genome stabilization at stalled replication forks. Genes Dev. 18: 1886-1897 (2004).
Hishida, T., Han, Y-W., Fujimoto, S., Iwasaki, H. and Shinagawa, H.
Direct evidence that a conserved arginine in RuvB AAA+ ATPase acts as an allosteric effector for the ATPase activity of the adjacent subunit in a hexamer. Proc. Natl. Acad. Sci. USA 101: 9573-9577 (2004).
Hishida, T.., Ohno, T., Iwasaki, H. and Shinagawa, H.
Saccharomyces cerevisiae MGS1 is essential in strains deficient in the RAD6-dependent DNA damage tolerance pathway. EMBO J. 21: 2019-2029 (2002).
Hishida, T., Iwasaki, H., Ohno, T., Morishita, T. and Shinagawa, H.
A yeast gene, MGS1, encoding a DNA-dependent AAA(+) ATPase is required to maintain genome stability. Proc. Natl. Acad. Sci. USA 98: 8283-8289 (2001).
プロフィール/メンバー
教授

菱田 卓
1992年 早稲田大学教育学部理学科(生物学専修)卒業
2000年 大阪大学医学系研究科博士課程終了(医学博士)
教授経歴
1992年 早稲田大学教育学部理学科(生物学専修)卒業
民間会社勤務
2000年 大阪大学医学系研究科博士課程終了(医学博士)
2000~2001年 日本学術振興会特別研究員(PD)
2001~2005年 大阪大学微生物研究所助手
2005~2007年 大阪大学微生物研究所助教授
2007~2011年 大阪大学微生物研究所准教授
2007~2011年 大阪大学大学院理学系研究科准教授(兼任)
2011年~ 学習院大学理学部教授
(生命科学科、生命分子科学研究所)
助教
赤沼 元気
学生
- D1 野田 俊輔 Noda Shunsuke
- M1 島田 遼 Simada Ryo
- M1 松本 彩花 Matumoto Ayaka
- B4 池田 百合香 Ikeda Yurika
- B4 石成 若菜 Ishinari Wakana
- B4 鈴木 萌美 Suzuki Moemi
- B4 高橋 梨紗 Takahashi Risa
- B4 古澤 啓至 Furusawa Keishi
- B4 松田 明香里 Matsuda Akari